RNA上的m6A修饰受到了广泛的关注,其对于RNA的命运调控涉及其生物学功能的各个方面,包括RNA的剪接、转运、降解、翻译等过程,m6A主要由METTL3/METTL14复合物催化产生,目前的研究主要集中在METTL3/METTL14对于细胞质中mRNA的调控研究。近年来已有研究报道指出METTL3以及m6A修饰参与染色质的调控,但尚不清楚METTL14是否在染色质上存在独立于METTL3以及m6A修饰的调控功能。
2023年6月13日,来自复旦大学生物医学研究院的沈宏杰、徐文绮等联合牛津大学Ludwig肿瘤研究所的Yang Shi教授在Cell Reports杂志在线发表了METTL14 Regulates Chromatin Bivalent Domains in Mouse Embryonic Stem Cells的研究论文。该研究发现METTL14调控小鼠胚胎干细胞的染色质二价结构域修饰,并且该调控不依赖于METTL3以及m6A修饰。

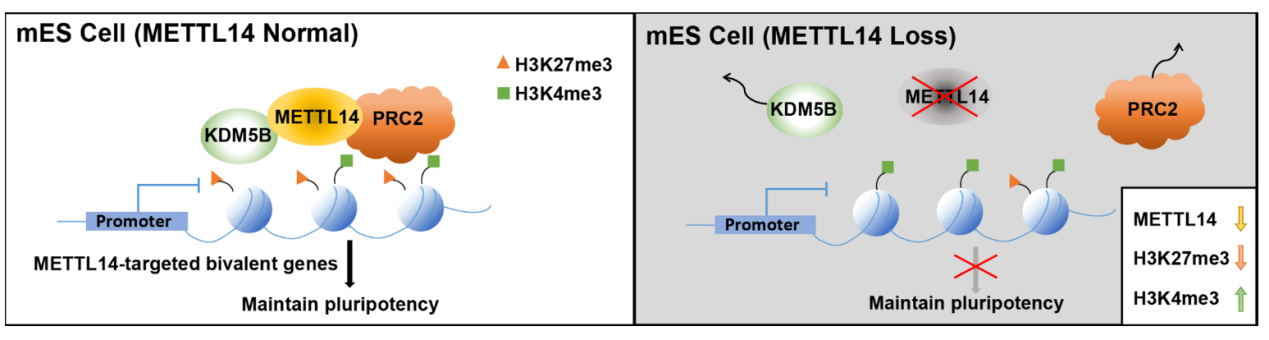
研究者发现在小鼠胚胎干细胞(mESC)中,METTL3和METTL14结合染色质的图谱有显著差异,不同于METTL3主要结合在H3K9me3修饰标记的逆转录转座元件IAP位点,METTL14主要结合在H3K27me3和H3K4me3修饰共同标记的二价结构域。并且METTL14缺失会导致H3K27me3的减少和H3K4me3的增加,从而导致二价结构基因激活。研究者进一步通过回补野生型以及与m6A功能相关的位点突变的METTL14蛋白(R298A、D312A)都能够回补H3K27me3和H3K4me3的变化,提示METTL14对于二价结构域上组蛋白修饰的调节是独立于METTL3介导的m6A功能的。在分子机制上,研究者发现METTL14能和H3K27me3的甲基转移酶PRC2和H3K4me3的去甲基转移酶KDM5B相互作用,并且敲除METTL14会导致PRC2和KDM5B在二价结构域上的结合降低,提示METTL14是通过促进PRC2和KDM5B在二价结构域上的结合从而影响组蛋白修饰,从而调控二价结构基因表达。
这项研究与今年2月份北京大学刘君团队和美国芝加哥大学何川教授团队合作在Protein & Cell上发表的文章METTL14 is a chromatin regulator independent of its RNA N6-methyladenosine methyltransferase activity具有一致性,两个研究都发现METTL3和METTL14在小鼠胚胎干细胞具有不同的染色质调控功能。但两者也存在不同,刘君、何川教授合作团队的研究主要关注H3K27me3修饰,并且发现METTL14缺失以后H3K27me3修饰上升,其分子机制可能通过METTL14招募H3K273去甲基化酶KDM6B实现。
由于METTL14缺失会导致小鼠胚胎干细胞状态的改变,在本项研究中,研究者构建了METTL14的快速降解细胞,利用dTAG-13小分子处理24小时快速降解内源METTL14蛋白(此时小鼠胚胎干细胞状态基本不受影响),此时H3K4me3修饰上升,但H3K27me3修饰只有比较微弱的降低。由于异染色质修饰H3K27me3相对稳定,H3K27me3可能需要比较长的时间才能降低,研究者利用EZH2的抑制剂处理去除细胞内的H3K27me3修饰后撤除抑制剂观察H3K27me3重建,同时利用快速降解系统降解METTL14蛋白,结果显示METTL14蛋白确实能够促进H3K27me3修饰的重建。
由于小鼠胚胎干细胞在分化过程中,存在染色质状态的改变,因此METTL14在小鼠胚胎干细胞不同的阶段是否存在不同的调控模式,还有待进一步探索。
复旦大学生物医学研究院牧蔓狄博士生是本文的第一作者,复旦大学生物医学研究院的沈宏杰、徐文绮、牛津大学Ludwig肿瘤研究所的Yang Shi为论文的通讯作者。
原文链接:https://doi.org/10.1016/j.celrep.2023.112650