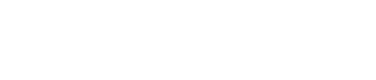锌离子(Zn2+)是人体内含量第二丰富的过渡金属离子,在细胞生长、增殖以及神经调节等多种生理过程中发挥重要作用。在特定脑区,突触释放的锌离子通过调控多种配体门控与电压门控离子通道,对维持正常的神经元回路功能起着重要作用,其稳态失衡与帕金森病、阿尔茨海默病、癫痫等多种神经系统疾病密切相关。Cys-环受体(CLR)超家族是一类五聚体配体门控离子通道,介导乙酰胆碱、5-羟色胺、GABA和甘氨酸等经典神经递质的快速突触传递。锌激活通道(ZAC)是CLR超家族中一个独特且尚未被充分研究的成员,与其他家族成员序列同源性较低(<20%),生理和病理意义尚不明确。ZAC蛋白可分布于人胎儿期大脑、脊髓、胰腺、胎盘等部位,在成年个体中则集中表达于参与认知、意识和情绪等功能的海马区、纹状体、杏仁核及丘脑。由于ZAC在大鼠、小鼠模式动物中不表达,限制了对其的生物学功能研究。针对常见的锌配位残基(如组氨酸、半胱氨酸、谷氨酸和天冬氨酸)进行的广泛突变,未能显著影响Zn2+诱导的ZAC信号传导。因此,ZAC如何受Zn2+调控的机制仍不清晰。
2026年3月31日,复旦大学生物医学研究院/附属眼耳鼻喉科医院屈前辉课题组与哥本哈根大学Anders A. Jensen教授、中科院物理所朱洪涛研究员合作,在Cell Discovery上发表了题为“Structural basis of human zinc-activated channel (ZAC) signaling and modulation”的研究论文,首次系统解析了人源ZAC在静息、锌离子激活及小分子抑制状态下的高分辨率结构,为理解这一非典型CLR受体的功能奠定了重要基础。

研究团队通过冷冻电镜技术,成功捕捉ZAC在无配体、部分及完全锌离子结合状态下的三维结构,并进一步解析其与两种拮抗剂结合的复合物结构。研究发现,ZAC亚基胞外结构域界面氨基酸Phe67、Arg88、Tyr176和Glu194可预形成一个弱相互作用网络,而Zn2+通过与芳香残基Phe67和Tyr176 形成“阳离子-π”作用,增强ZAC通道的电导性。电生理功能实验表明,破坏该相互作用显著阻断锌离子对ZAC的激活程度(图1)。同时F67W、Y176W突变体保留对Zn2+响应效果,揭示了该作用网络中芳香残基的可替代性。
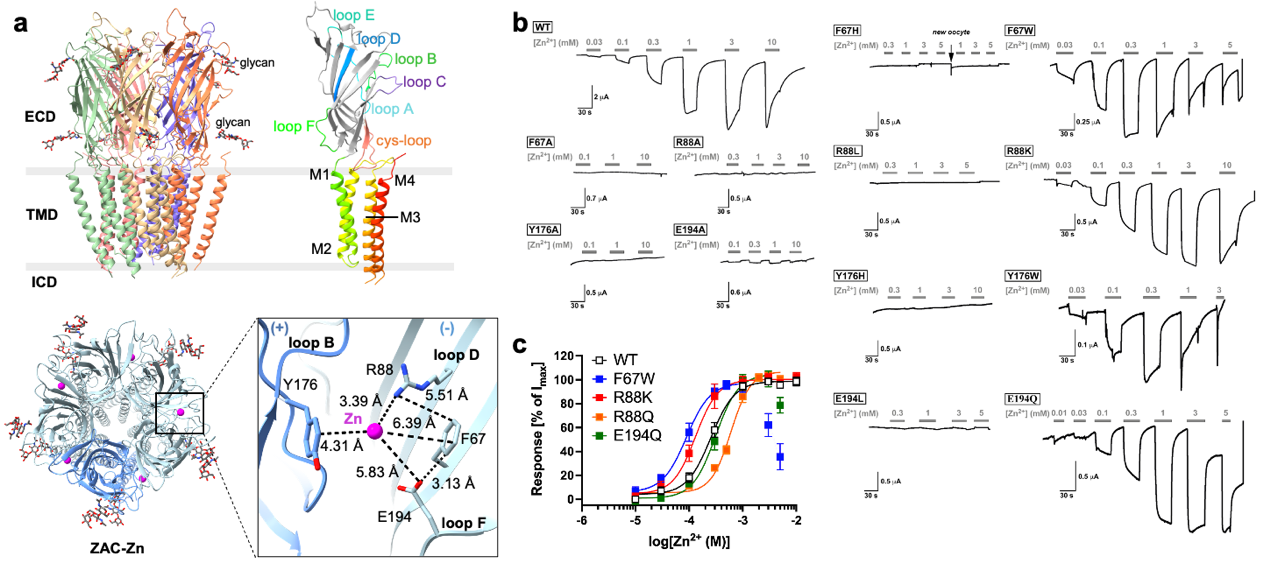
图1. 人源ZAC的ECD区域Zn2+-π互作网络分析。a,ZAC结合Zn2+的五聚体结构及ECD结合位点的细节展示。b,采用双电极电压钳记录技术,检测在表达野生型ZAC及突变体的非洲爪蟾卵母细胞中,Zn²⁺诱发电流响应结果。c,野生型及突变体F67W、R88K、E194Q ZAC的激动剂浓度-响应关系曲线。
此外,研究还揭示了两种拮抗剂的抑制机制:选择性拮抗剂TTFB通过嵌入跨膜区直接阻塞离子通道;而非选择性拮抗剂d-筒箭毒碱则采用“双位点”结合模式,既竞争锌离子结合界面,也进入通道孔内部,从而高效抑制通道活性 (图2)。
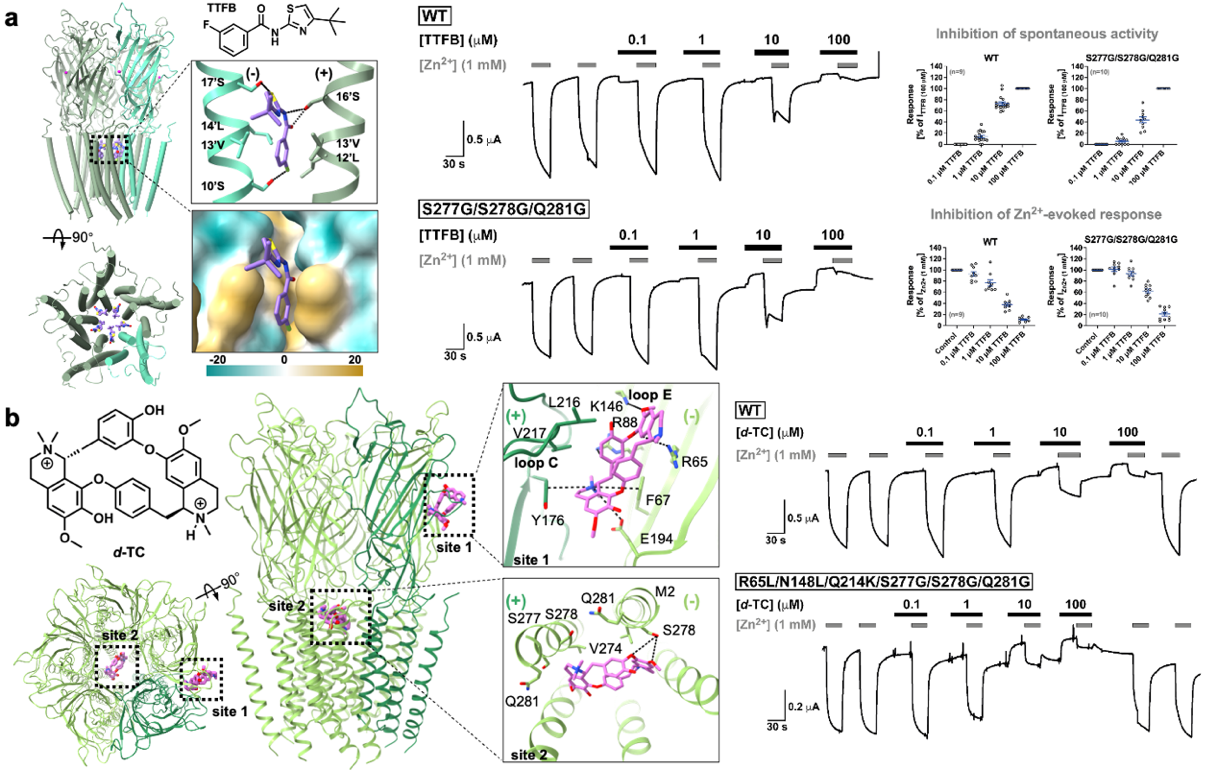
图2. 拮抗剂TTFB和d-筒箭毒碱(d-TC)抑制ZAC的分子机制。a,TTFB(紫色)结合在ZAC跨膜区的相邻M2螺旋之间,形成通道孔道阻滞。b,d-TC在ZAC中存在两个结合位点:位点1(ECD亚基界面)和位点2(通道孔道入口)。
研究还聚焦于人群中高频存在的功能变异体ZACA¹⁵²,从结构上阐明其功能减弱的机制:该变异破坏了原本由T152构建的亚基间关键相互作用网络,影响信号从结合位点向通道门的传递效率(图3)。
图3. ZACA152变异体功能减弱的分子基础及离子通透路径。a,野生型ZAC(T152)与变异体ZACA152在关键相互作用网络(Thr152/Leu126)上的结构差异。b,T152支点网络突变体的电生理结果。
这项工作不仅为理解ZAC这一非典型受体提供了关键结构基础,揭示了其独特的激活与抑制机制,也为未来针对ZAC的相关药物研发提供了重要的理论依据和结构蓝图。
该工作主要由复旦大学附属眼耳鼻喉科医院助理研究员周子璇及哥本哈根大学Anders A. Jensen教授共同完成,复旦大学生物医学研究院龙永惠、晁钰琳、杨传辉等参与工作,并得到了复旦大学附属眼耳鼻喉科医院舒易来教授、中国科学院物理研究所朱洪涛研究员、复旦大学脑科学研究院唐逸泉研究员等的重要支持。研究得到了国家自然科学基金、国家重点研发计划、非传染性慢性病-国家科技重大专项、丹麦独立研究理事会医学科学部、隆德贝克基金会以及中国博士后科学基金等多个项目的支持。
原文链接:https://www.nature.com/articles/s41421-026-00878-5